It has long been known that lake levels in the American south west have fluctuated over a millennial timescale. Identification of ancient lake shoreline features began with early explorers of the continent. The first detailed studies were conducted by pioneering American geologists such as G.K. Gilbert and I.C. Russell in the late 1800s, who studied Lake Bonneville, now the remnant Great Salt Lake in Utah, and Lake Lahonton, located in northwestern Nevada. After 1900 many other dry lakes came under scientific investigation.
Lake Estancia in New Mexico, was one of the first studied in 1903. In 1933 a sediment core was drilled, which reached 600 feet in length, giving one of the longest sedimentary deposit records in America. Work continued for decades at this lake with findings helping to elucidate palaeoclimate swings and providing fossils such as Mammoth, which could be radiocarbon dated to build a chronology of wet and dry phases.
It was also discovered that some dry salt, lakes such as Lake Otero, also in New Mexico, preserved footprints of various animal species on their surfaces. These included Mammoth; Giant Ground Sloth; Camel, Bovids (probably Bison); Dire Wolf; American Lion; other smaller felids, and Sabre Tooth Cat.
Study of these footprints and sediment record at Lake Otero and the adjacent Lucero has provided a wealth of information about Pleistocene animal distribution, locomotion and behaviour.

Study sites mentioned in this blog post, adapted from Bustos et al. (2018).
Below are some of the remarkable foot/paw prints, of various Pleistocene animals discovered at White Sands:

Eroded and raised Mammoth tracks from the White Sands National Monument. Image from Lucas (2007). Original caption reads: Figure 3. Selected mammoth tracks from the southern shore of Lake Lucero. Arrows indicate inferred direction of travel.

Mammoth foot forming tracks, or “Walking in Mud” as Bennet et al. (2019) describe it.

Mammoth, human and sloth tracks, preserved together for eternity (or until erosion takes them) in the sediments of White Sands National Monument. Picture credit: Bennett et al. (2020).
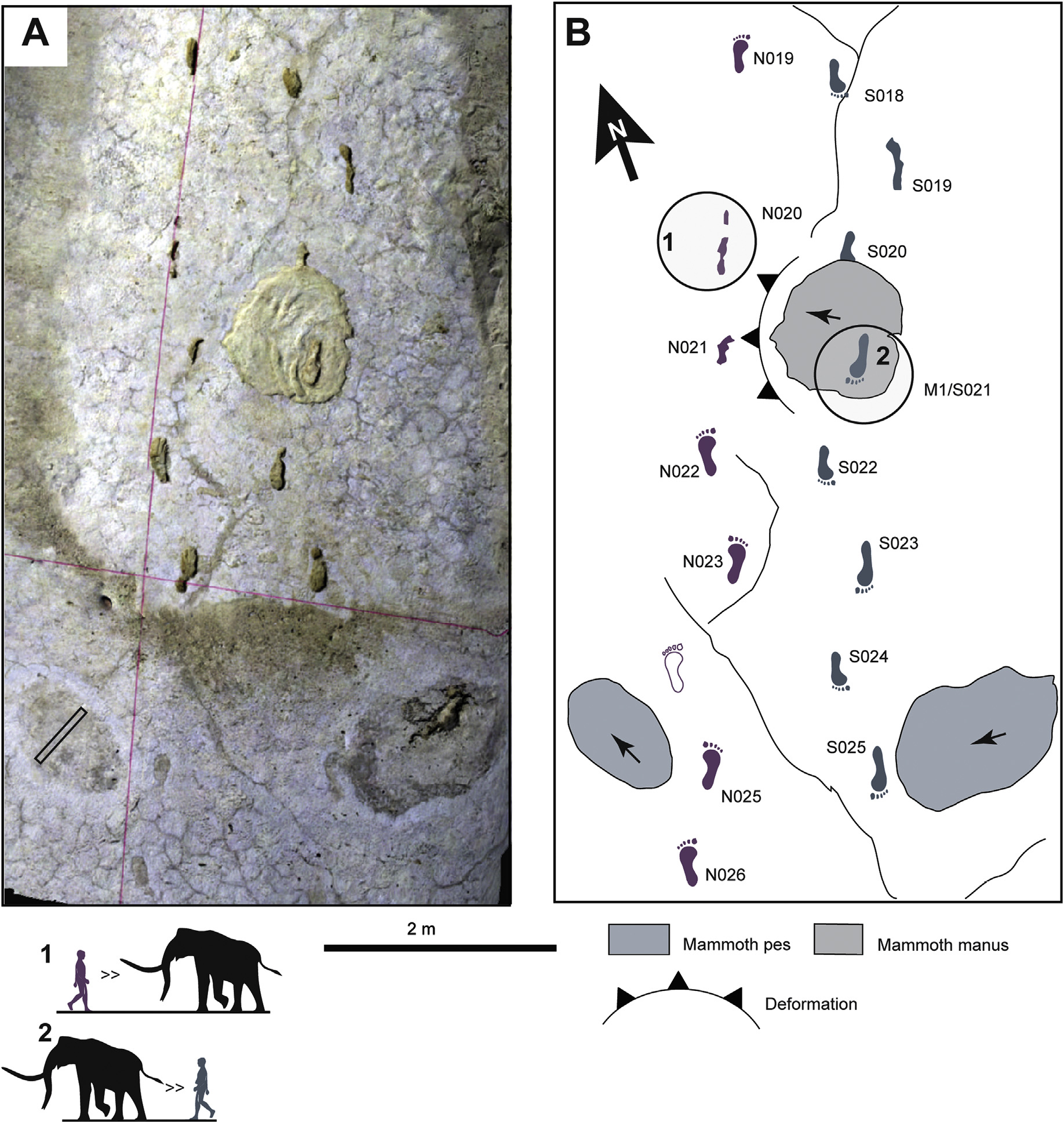
Interaction between a Mammoth and Human at White sands, from Bennet et al. (2020). In case you wondered, the pes is the rear foot, while the manus is the forefoot.

Mammoth trackway, eroding out from Santucci et al. (2014).

Track depth images in false colour, from a 3D ground penetrating radar study by Urban et al. (2019). Original caption reads: Figure 3. (a) The principal tracks and trackways observed at the study site which are split into Location-1 and Location-2 (shown in true spatial relationship). (b) GPR amplitude slice (2.0 to 4.0 ns). Human prints that were excavated and used for analysis are indicated with (+) while an unexcavated sloth trackway (identified in subsequent fieldwork) is indicated with (x).

Giant Sloth tracks, colour rendered in 3D, adapted from Bennett et al. (2020).

Human footprint superimposed on Giant Sloth footprint. Is this human stalking this sloth? Adapted from Bustos et al. Fig. 2 (2018).

Further along the Sloth/Human trackway a unique event is captured in the fossilised sediments: the sloth appears to rear on its hind legs and rapidly perform pirouettes. From Bustos et al. (2018). Original caption reads: Flailing circle made by a sloth reaching forward with its forelimbs and leaving knuckle and claw impressions.
In the accompanying text, Bustos, et al. (2018) describe the interactions between the sloth and human(s) present: “The sloth trackways show several circular (and elliptical) track patterns that have never been reported. They consist of one or more deformed pes tracks surrounded by a circle of irregular impressions, which show evidence of manus claw marks. The manus impressions are distinctive, 120 to 140 mm long, with pointed termini at both the distal and proximal ends, showing evidence of one or more claw scrapes. The pes impressions at the center of these circular patterns are irregular and in some cases circular, and they appear to represent heel or pivot traces. In one case, a line of human toe impressions leads to the circle center, suggesting that someone approached on raised toes. In another, an adult human track occurs in the middle of one circle. These structures occur both independently and in association with overlapped human-sloth trackways. Sloth trackways show sharp changes in the direction of travel. These inflections are associated with concentrations of human tracks. In the absence of human tracks, sloth trackways progress in a straight or curvilinear fashion. Measures of tortuosity confirm this. We have evidence for: (i) sloth and human tracks in close spatial association, (ii) superimposed human and sloth tracks with geological tracks were open and unfilled with water and/or sediment when the human trackmaker followed, and the alignment was perfect. Pore-water conditions demonstrate constancy during both track-forming episodes, evidenced by the deformation structures present. In terms of the second test, the tortuosity of sloth trackways increases when human tracks are present, with sharp direction changes that suggest evasion. In addition, the circular sloth trackways are consistent with defensive behaviors in which sloths reared on their hindlimbs, freeing their forelimbs for defense. We termed these structures “flailing circles.” To be clear, the human-sloth interactions are not limited to one sloth trackway (or track size) but to several. It is, however, difficult to say definitively whether the sloths were traveling as a group and therefore were being collectively harassed or whether these features represent successive harassment events. The co-location of these trackways favors the former, in which case the results have implications for the social behavior of sloths. We argue that the tracks evidence temporal and spatial associations of sloths and humans and infer that humans actively stalked and/or harassed sloths, if not hunted them.”
Bustos et al. (2016) also give a probable sequence of events for Palaeolake Otero and a window of time for the Sloth Human interaction:


Bennett et al. (2020) also report another Sloth/Human interaction. This time, a double human trackway in association with that of a sloth. Bennett offers a slightly opaque explanation of what went on: “..the two human trackways time-bracket the passage of the mega-fauna; north-bound is crosscut by the sloth tracks, south-bound cuts the sloth tracks. The ground sloth approaches from the east, perpendicular to the human trackway and does a circular shuffling ‘dance’ over the north-bound trackway. The ground sloth tracks are distinctive and have a more plantigrade form than those reported by Bustos et al. (2018) from the west side of Alkali Flat. Overprinting of the manus by the pes is present, with sharp curved extensions to some prints and a manus impression is distinct close to the north side of the circular dance. The lack of manus impressions and the plantigrade nature of the tracks compared to those described elsewhere (McDonald, 2007; Melchor et al., 2015) may indicate that at least initially, the giant ground sloth rose onto it’s hind legs, perhaps to scent or scout for the human, much as bears do today. The exact locomotion of the animal, however, during this shuffling circular-movement is not clear, but the double placement of tracks to create ‘wing-like’ impressions is distinctive and suggest small adjustments in foot placement.”
Incidentally, Bennett et al, (2020), go into some morphological detail to identify the most likely species of sloth that made the tracks: “Both Nothrotheriops and Paramylodon are known from body fossils in the vicinity of Palaeolake Otero and both have a pedolateral foot. However, Nothrotheriops has a higher arch with just the posterior part of the tubercalcis in contact with the ground while Paramylodon has a lower arch with all of the tuber calcis in contact with the ground. This might suggest that the track-maker was more likely Paramylodon, notwithstanding potential effects due a more upright posture.”

Line drawing of the pes (rear foot) of the likely species of sloth which made the tracks at White Sands. Image credit: McDonald (2007). Original caption reads: Figure 2 Comparison of the pes of the four different groups of ground sloths in which the hind foot is rotated in the pedolateral position. A. Mylodontine. Paramylodon harlant..
Tracks of a number, of types of other Pleistocene animals have been found at White Sands. These include both herbivores and carnivores.

Left to right above mammoth, camel, saber tooth cat, and dire wolf. Image montage from Bustos (2017).

Large Pleistocene camel track eroded out and standing proud of the surface at White Sands. The track was photographed in 1981, by Robert A. Corley (US Army) who was accompanying a group of scientists, as their official escort onto the White Sands Missile Range. The group included Spencer Lucas, John Hawley, David Wolberg and David Love. The group examined the footprints and identified them as belonging to Colombian Mammoth and a camel species. From Lucas et al. (2002).
Most interestingly, Lucas et al (2002) comment: “Tracks of proboscideans and camels in the Neogene mammal footprint record. Body fossils of camel (Camelops sp.) and mammoth (Mammathus columbi) are found in the Otero Formation, and these are the probable trackmakers of the mammoth and camel tracks at NMMNH locality 4979.
At the locality, all the preserved trackways are oriented nearly east-west. This suggests the animals walked to and from Lake Otero, most likely to drink. Although human artifacts (chipped stone) have been found near the tracksite, there are no human tracks and no demonstrable human association.”
Radiocarbon dates obtained after the publication of the Lucas et al. (2002) and by Allen et al. (2006) put some of the trackways as being formed prior to 30,000BP, with others dated between 14,000 and 23,000BP. This raises the possibility that man was in the region far earlier than believed.

Large carnivore track, probably American Lion (Panthera atrox) from Santucci (2014), note the roughly circular shape, reminiscent of modern African Lions.

Probable American Lion (Panthera atrox) from the NPS website (2022). Note that the print has only four toes, making this a rear paw print, also the claws are retracted, meaning the lion was walking and not about to strike prey. However, Childs (2014) attributes this track to the Short-Face Bear (Arctodus pristinus).


Top and above: Small carnivore tracks photographed at White Sands from Santucci et al. (2014).
The number and variety of tracks discovered at White Sands, increased over the years, with greater surveillance from scientists. However, due to their ephemeral nature, of many tracks and trackways, researchers began to seek new methods with which to gather the maximum amount of data from the constantly eroding tracks and to look deeper into the sediment layers to detect buried footprints.
Urban et al. (2019) reported: “How researchers detect and record fossil footprints is a burgeoning area of method development in contemporary ichnology, and crucial to both maximizing the information yielded and also preserving these fragile traces of the past. Former lake beds and playa sites, which occur extensively across the Americas and in parts of Africa, have the potential to hold these latent ichnological archives.
Here we present findings from White Sands National Monument (WHSA), New Mexico, USA. Our work demonstrates the effectiveness and efficiency of non-destructive GPR for detecting and documenting fossil footprints in soft sediments, including human tracks.” and “Importantly, the survey was able to resolve targets for excavation that are not visible at the surface.”
This proof of concept led Bennett et al. (2021) to actively seek older deposits, in which human footprints were extant: “The site at White Sands National Park (WHSA) Locality 2 was selected for excavation as result of (a) an initial ground-penetrating-radar survey by T.M.U. that revealed potential targets following the method outlined in Urban et al. (2019); (b) field prospecting over several years by D.B. that revealed potential tracks at the surface by color and textural contrasts; (c) preliminary 14C ages that established the general age of the deposits based on samples collected in April 2019 by D.B., K.B.S., and J.S.P. along a transect ~50 m north of WHSA Locality 2; and (4) a preliminary surface excavation in September 2019 conducted by D.B., P.J.M., C.C., and M.R.B.”
The method worked like a dream: Bennett and his team found human footprints throughout the depth of the profile they cut with a mini-digger. Tracks of mammoth and canid were also discovered.
Bennett et al. (2021) give an overview of their findings: “WHSA Locality 2 is located on the eastern side of Alkali Flat on a shallow (<6 m) erosional scarp formed between the current playa and the White Sands gypsum dune field. A sedimentary sequence exposed by trenching consists of 1.25 m of lacustrine clays and silts intercalated with thinly bedded gypsiferous and siliciclastic sands, silts, and clays, which represent a transition from a lacustrine ecosystem to an alluvial regime in response to changing hydroclimate conditions during the late Pleistocene (Figs. 2 and S2). Endocarps of ditch grass (Ruppia cirrhosa) occur as discontinuous layers throughout the sequence.
Footprints (humans, proboscidean, and canids) occur at all levels both in cross section and on excavated surfaces adjacent to the trench (Figs. 1B-E, 2, S4-S11). A total of 61 human tracks have been identified and documented.
Human footprints reported from all surfaces have good anatomical definition (i.e., visible heel impressions, medial longitudinal arches, and toe pads) consistent with modern humans..”

Adapted from Bennett et al. (2021) supplementary materials S5. Original caption reads: “Figure S5: Tracks on TH1. A. Surfaced 3D model of TH1 located in the bottom of the north-south side trench. B. General view of the north-south side trench and the TH1 surface. C. Surfaced 3D model of the southern end of the north-south side trench. Note the gently deformed TH2 surface, deformed by an overlying proboscidean track. The dark layer is a seed layer.


Top: selection of human footprints excavated by Bennett et al. (2021) at White Sands. Adapted from their Fig. 1. Original caption for B, C and E reads: Figure 1: B. Human footprints on track horizon 4 (TH4). C. Human footprint on track horizon 5 (TH5) located in the base of the main trench. E. Human trackways on surface TH4.
Above : A wider view of the trackways on surface TH4, from Danielle (2021).
The seed layers of ditch grass (Ruppia cirrhosa) found during the course of the excavation, were to prove crucial. Radiocarbon dating established a chronology for these layers and hence the dates the human footprints were impressed into the surface. Bennett et al. (2021) describe their findings: “We established chronological control for the footprint-bearing sediments using radiocarbon dating of in situ layers of macroscopic seeds of the aquatic plant Ruppia cirrhosa. Seed layers were sampled from throughout the sedimentary sequence and yielded calibrated ages that range from 22.86 ± 0.32 to 21.13 ± 0.25 ka (N=11) and maintain stratigraphic order within the uncertainties between TH2 and TH6.”
The authors considered whether the hard water (reservoir) effect could have made the radiocarbon ages too old, which would be the key point of contention after publication.
They summarise their reasoning for believing that their radiocarbon ages to be correct by using three lines of evidence: “Radiocarbon dating of aquatic material, such as R. cirrhosa, may be subject to hard-water (or reservoir) effects that could conceivably make these ages too old, so we explored the potential for these effects at WHSA Locality 2 using three lines of evidence.
First, the geologic and hydrologic settings along the shallow lake margin that hosts the human trackways make it unlikely that stands of emergent aquatic plants would harbor significant hard-water effects.
Second, our calibrated 14C ages maintain stratigraphic order even when samples were separated by centimeter(s), which would not be the case if hard-water effects were large and variable.
Finally, we evaluated independent chronologic data from the Tularosa Basin and found that terrestrial and aquatic material yielded concordant 14C ages between ~44 and 25 ka, which demonstrates that hard-water effects in Paleolake Otero were less than a few hundred years during this period. In our view, an improbable series of events would be required to introduce a large hard-water effect by ~23 ka when for the previous ~20 kyr they were minimal. For these reasons, we conclude our radiocarbon-based chronology is robust.”
A statistical analysis of the distribution of the C14 ages was also carried out and confirmed the above.

The deepest and oldest footprints from the base of the main trench. These are part of track horizon 5 (TH5) and dated to ≈ 23,000BP. Image credit: Schuster (2021).
The authors also attempted to independently verify the chronology of the deposits using Uranium-series dating. This was unsuccessful due to a failure in the process. Bennett et al. (2021) in their supplementary materials say: “After this second digestion step was complete, significant undissolved residue remained. After inspection under the binocular microscope, this residue was clearly dominated by undissolved gypsum. Total digestion of samples was not obtained; therefore, apparent U and Th concentrations are inaccurate and not reported.”

Bennett et al. (2021) composite stratigraphy. Original caption reads: “Figure 2: A. Composite stratigraphy, calibrated ages, and dominant hydrologic regime for sediments exposed in the trench at WHSA Locality 2. Filled circles denote 14C samples; filled square denotes uranium-series sample. The calibrated 14C ages were used to construct the age models shown in Figure 3. B. Summary of footprint evidence on each horizon [number of human tracks, number of trackways, minimum number of track makers (MNT), track-maker age, stature, and fauna].
Conclusions
While Bennett et al. (2021), have taken steps to ensure the accuracy of their C14 measurements of the footprint bearing trackways at the WHSA locality 2, and to my untrained eye seem to advance a very plausible argument for man in America ≈ 23,000BP, others disagree.
Madsen et al. (2022) question the data, on the very grounds that Bennett et al. identified as possible grounds of inaccuracy. Madsen attacks the Bennett interpretation of the reservoir effect as of only being of the magnitude of a few hundred years. Instead Madsen et al. (2002) posit that the reservoir effect could have made the Ruppia seeds appear 1000’s of years older than they actually are.
In response Pigati et al. (2022) more than adequately dispose of the Madsen et al. (2022) arguments. In particular they show how Madsen et al. (2022) us the wrong comparative data to undermine their results. To me, this ends the argument in favour of the original Bennett et el paper (2021).
Where does this leave us? One can conclude:
The footprint dates are accurate, and therefore, man entered the Americas some time before 23,000BP.
As Bennett et al. (2021) theorise: “The evidence presented here confirms that humans were present in North America before the glacial advances of the LGM closed the Ice-Free Corridor (9, 26) and Pacific Coastal Route to human migration from Asia (7). The overlap of humans and megafauna for at least two millennia during this time suggests that if people were hunting megafauna, the practices at least initially were sustainable.”
This one of the first times, I have seen the idea of a use of the ice-free corridor being used prior to its closing by the LGM. This is a bold statement and an idea that has long waited for a discussion in print. Bravo to Bennett et al. (2022).
Then there are the chipped stones. Those discovered by Lucas et al. (2002) at NMMNH locality 4979 were near tracks of mammoth and camel travelling east-west to drink at the lake. One can imagine ancient people biding their time in the ditch grass thickets, waiting for suitable animals to pass. As they waited they filled their time making spear points with which to bring down prey. Allen (2006) dated some of the tracks to before 30,000BP! Bennett and David Bustos are still working at White Sands, who knows what discoveries they will make in the next few years? Maybe they will confirm the 50,000BP+ dates of human occupation obtained by MacNeish and Libby (2003) at the nearby Pendejo Cave?
References
Allen, B.D., Love, D.W. and Myers, R.G., 2006. Preliminary age of mammal footprints in Pleistocene lake-margin sediments of the Tularosa Basin, south-central New Mexico. New Mexico Geology, 28, pp.61-62.
Bennett, M.R., Bustos, D., Belvedere, M., Martinez, P., Reynolds, S.C. and Urban, T., (2019). Soft-sediment deformation below mammoth tracks at White Sands National Monument (New Mexico) with implications for biomechanical inferences from tracks. Palaeogeography, Palaeoclimatology, Palaeoecology, 527, pp.25-38.
Bennett, M.R., Bustos, D., Odess, D., Urban, T.M., Lallensack, J.N., Budka, M., Santucci, V.L., Martinez, P., Wiseman, A.L. and Reynolds, S.C., (2020). Walking in mud: remarkable Pleistocene human trackways from white sands national park (New Mexico). Quaternary Science Reviews, 249, p.106610.
Bennett, M.R., Bustos, D., Pigati, J.S., Springer, K.B., Urban, T.M., Holliday, V.T., Reynolds, S.C., Budka, M., Honke, J.S., Hudson, A.M. and Fenerty, B., (2021). Evidence of humans in North America during the last glacial maximum. Science, 373 (6562), pp.1528-1531.
Bustos, D., (2017). Much More than a Sand Box: Fossil Tracks from the Lakes of the World’s Largest Gypsum Dune Field. Park Paleontology News - Vol. 09, No. 2, Fall 2017. At: https://www.nps.gov/articles/park-paleo-fall-2017-bustos-whsa.htm accessed 01/05/2022
Bustos, D., Jakeway, J., Urban, T.M., Holliday, V.T., Fenerty, B., Raichlen, D.A., Budka, M., Reynolds, S.C., Allen, B.D., Love, D.W. and Santucci, V.L., (2018). Footprints preserve terminal Pleistocene hunt? Human-sloth interactions in North America. Science Advances, 4(4).
Childs, C. (2014). Seeing Mammoths, at https://www.lastwordonnothing.com/2014/10/28/seeing-mammoths/ accessed 02/05/2022
Danielle, M. (2021) Ancient footprints reshape scientists' understanding of early life in Americas. Accuwether at: https://www.accuweather.com/en/weather-news/ancient-footprints-white-sands-new-mexico-23000-years-old/1023896 accessed 17/04/2022
Lucas, S.G., Morgan, G.S., Hawley, J.W., Love, D.W. And Myers, R.G., (2002). Mammal Footprints from the Upper Pleistocene of the Tularosa Basin, Dona Ana County, New Mexico, in: Geology of White Sands, Lueth, Virgil; Giles, Katherine A.; Lucas, Spencer G.; Kues, Barry S.; Myers, Robert G.; Ulmer-Scholle, Dana; [eds.], New Mexico Geological Society 53rd Annual Fall Field Conference Guidebook, 362 p.
Lucas, S.G., Allen, B.D., Morgan, G.S., Myers, R.G., Love, D.W. and Bustos, D.A.V.I.D., (2007). Mammoth footprints from the upper Pleistocene of the Tularosa Basin, Doña Ana County, New Mexico. Cenozoic Vertebrate Tracks and Traces, 42, pp.149-154.
MacNeish, R.S. and Liddy, J.G. eds., 2003. Pendejo Cave. UNM Press.
Madsen, D.B., Davis, L.G., Rhode, D. and Oviatt, C.G., (2022). Comment on “Evidence of humans in North America during the Last Glacial Maximum”. Science, 375(6577).
McDonald, H. G., (2007). Biomechanical Inferences of Locomotion in Ground Sloths: Integrating Morphological and Track Data. Cenozoic Vertebrate Tracks and Traces: New Mexico Mus. Nat. Hist. Sci. Bull. 42, 201-208.
NPS (2022). American Lions, at: https://www.nps.gov/whsa/learn/nature/american-lions.htm accessed 02/05/2022
Pigati, J.S., Springer, K.B., Bennett, M.R., Bustos, D., Urban, T.M., Holliday, V.T., Reynolds, S.C. and Odess, D., 2022. Response to Comment on “Evidence of humans in North America during the Last Glacial Maximum”. Science, 375(6577).
Santucci, V.L., Tweet, J., Bustos, D., Von Haden, J.I.M. And Varela, P., (2014). Vertebrate Paleontological Resources from National Park Service Areas In New Mexico. Vertebrate Paleontology in New Mexico: Bulletin 64, p.41.
Schuster, R. (2021). Footprints Prove That in Ice Age, Humans Were Already in America. Haaretz, at: https://www.haaretz.com/archaeology/.premium-clovis-debunked-footprints-show-humans-in-america-23-000-years-ago-1.10235486 accessed 17/04/2022
Urban, T.M., Bennett, M.R., Bustos, D., Manning, S.W., Reynolds, S.C., Belvedere, M., Odess, D. and Santucci, V.L., (2019). 3-D radar imaging unlocks the untapped behavioral and biomechanical archive of Pleistocene ghost tracks. Scientific Reports, 9(1), pp.1-10.
Very interesting post!. On the subject of it, I have recently found an nice paper, Bente Philippsen (2013), about the nature of the so called “reservoir effects” that affect C14 measurements... that among other considerations, also analyses the particular case of an aquatic plant; and more precisely how can it obtain its carbon from sources with lower C14 than that of the atmosphere, thus leading to an apparently older age. Although this author reports exceptionally high values of 2700 years AP for some hardwater Estonian lakes, it can be inferred from this work that “average” reservoir effects of submerged plants that live in mixohaline waters (like Ruppia cirrhosa) can range (depending on the sources of carbon which they has been interacting with) between a few hundred of years up to as high as 2000 years AP (understand; not much more than this…), thus suggesting, at least, that the claims from Madsen et al. (2022) about inaccuracies of the C14 ages obtained by Bennet et al. (2021) due to high values of reservoir effects… even when these biases could have been possible (??), they could hardly have been large enough by themselves to invalidate the presumption of human presence in the White Sands before the LGM´s peak (that occurred around 18/20 Ka or so), given the C14 obtained ages from 21 to 23 Ka…
ReplyDeleteWhether these people had just arrived somewhat before those times via coastal route, or simply they were in this continent since much more ancient times (there are not so weak signs of this…), I would totally agree with you in that this is an absolutely relevant evidence, that certainly adds to the already overwhelming list of ones indicating pre-LGM human presence in America…
Best regards
Marcelo
Philippsen, B. The freshwater reservoir effect in radiocarbon dating. herit sci 1, 24 (2013). https://doi.org/10.1186/2050-7445-1-24