Discovered in 1848 by Captain Edmund Flint, or so the story goes. Flint, a British Royal Navy officer was overseeing the extraction of limestone to reinforce and rebuild the military fortress’s fortifications. The renovation of the stone walls had begun in 1841, after General Sir John Thomas Jones of the Royal Engineers conducted a study of Gibraltar's defenses and recommended improvements.
The stone for the
construction of Jones’ "retired batteries" was got, by the use of
gunpowder in quarries such as Forbes’ quarry. Why Capt. Flint, a naval officer
was overseeing the work and who the workers were, is unknown.
A local, likely apocryphal
story has it that, consequent to blasting Flint retired to a campaign chair in
the shade, while local Gibraltar men set about shifting the stone. Some while
later he was disturbed by a shout, put up by the workers: removal of blocks had
revealed a previously hidden cave without any apparent external openings. The
workers had looked in and apparently seen bones. This is what caused them to
send up alarums. Flint it seems, availed himself of a skull found in the cave.

Gibraltar 1 cranium, a female and probably between 60,000 to 120,000 years old. Image credit: Pavid (2019).
At least the part of the story about the skull being found in a previously sealed cave was current among the learned men of the time. This is proved by the quip of George Busk (1864): “..in many respects, it [Gibraltar 1] is of infinitely higher value than that much-disputed relic, Neanderthal 1… [It] adds immensely to the scientific value of the Neanderthal specimen, if only as showing that the latter does not represent, as many have hitherto supposed, a mere individual peculiarity, but that it may have been characteristic of a race extending from the Rhine to the Pillars of Hercules; for, whatever may have been the case on the banks of the Dussel, even Professor Mayer will hardly suppose that a rickety Cossack engaged in the campaign of 1814 had crept into a sealed fissure in the Rock of Gibraltar.” Here Busk is referring to the theory of Mayer that Neanderthal 1 was a rickety Cossack. Mayer’s published views are well summarized by Schrenk and Müller (2010): “He confirmed the Neanderthaler's "rickety" changes in bone development... Mayer argued, among other things, that the thigh and pelvic bones of the Neanderthal man were shaped like those of someone who had spent all his life on horseback. The broken right arm of the individual had only healed very badly and resulting permanent worry lines due to the pain were the reason for the distinguished brow ridges. The skeleton was, he speculated, that of a mounted Russian Cossack, who had roamed the region in 1813/14 during the turmoils of the wars of liberation from Napoleon."
While Busk was humorously dismissive of Mayer’s interpretation of the other Neanderthal skull and remains, obviously something about the circumstances of the discovery, of Gibraltar 1, didn’t sit well with him. Undoubtedly, it was the condition of the Gibraltar 1 skull, with its sandy lime concretions that caused his disquiet. Busk, therefore determined to visit the find spot himself. Menez (2018), explains the circumstances of his subsequent exploration at Gibraltar: “Busk had planned to visit Gibraltar, noting in The Reader (Busk 1864a) that the purpose, with the full support of the Governor, Falconer and Busk (1865), was “to examine on the spot the conditions under which the different classes of fossils occur in the caverns &c”. The trip would wait, however, until after the Bath [BAAS] meeting, perhaps becoming even more important than hitherto after it became apparent that information related to the skull, which had engendered interesting debate in Bath, was scarce (Busk 1864b), Busk noting that: “The only information which we have been furnished respecting the situation in which it was found, is to the effect that it was dug up in the course of some excavations being made in what is termed ‘Forbes’ Barrier’… The exact locality, however, does not appear to rest upon any very certain evidence, and it may perhaps turn out that this interesting relic was derived from some other situation in the rock.” Busk, accompanied by Hugh Falconer and the British physician and travel writer Henry Holland, arrived in Gibraltar aboard the Poonah on 24 September 1864.” Busk, Falconer and Lieutenant Alexander Burton-Brown, thereafter visited Forbes’ Quarry. Brown (1867) later reported:
“..in October, 1864, examining with Professor Busk the slope of the old quarry of Forbes [sic] we found the matrix in which we believed the skull had been imbedded, which was a raised beach of about 100 ft. above sea level, and which was seen everywhere to be cropping out of the limestone slope, the inner portion of the beach being covered with broken pieces of calcareous rock, and the outer being broken off, disjointed masses being carried down by the crumbling masses of limestone; many subsequent visits confirm me in the opinion of this being the veritable matrix.”
Busk (1865) reporting
his observations at Forbes’ Quarry said “…from the matrix with which it [the
skull] was thickly covered, and which contained a very large proportion of
coarse rolled siliceous sea-sand, similar to that which is blown up in such
large quantities against the north-eastern end of the rock, it was apparent
that it had been lodged in the superficial part of the talus in which the quarry
is worked.”

Top: Busk’s photographs of the skull before removal of the matrix. Bottom: Busk’s photographs of the skull following partial removal of the matrix. Bottom: the skull today. Picture credit: Archives of the Royal College of Surgeons.
In 1865, mid-way
through his description of Gibratar 1, Busk received an urgent message to
attend to his close friend Hugh Falconer. Falconer had started to feel feverish
on the 19th of January. According, to, Murchison (1868) “the attack
developed into acute rheumatism, complicated with disease of the heart and
lungs, which proved fatal.” Busk was at his side as he died. Without Falconer, Busk
apparently could not continue, and he never finished his analysis of Gibraltar
1.
Within a year of Busk’s announcement that the Forbes skull was an object of immense scientific importance, it had retreated back into the shadows once again. The skull had been found but then quickly lost, tucked into a back room of Busk’s Royal College of Surgeons, collecting dust.
Other scientists
visited the site of Forbes’ quarry in latter, years to try to further clarify
the exact find spot. Results were
varied, and somewhat confusing.
Samuel William Turner, Surgeon to the Colonial Hospital in Gibraltar, it seems was encouraged, by Sir Arthur Keith, to seek what information he could on the Forbes Quarry site. Turner (1910) reported back: “the sloping talus which abutts on the perpendicular line above of the north face of the Rock at this point forms with the limestone the quarry referred to, and there is a distinct line of demarcation between the limestone and the conglomerate of the talus.” He then conjectured a possibility that has been perpetuated in the published literature ever since, describing that there was a small cave at the juncture between the conglomerate and the limestone, Turner surmised that “possibly the skull may have been found in this cave”. His description of the cave: “40 ft long, 8 wide” and requiring him to “stoop somewhat”, it showed no trace of osseous remains. Turner (1910), wrote again to Keith on 28 April 1910, informing him that “I fear we shall not be able to definitely locate to within a few yards the spot where the skull was found, or to say whether it was found embedded in the conglomerate of the talus at Forbe’s [sic] Quarry, or lying on the floor of the cave which exists in that quarry.”
Duckworth (1911),
also visited the quarry in 1910, with William Turner, Colonel Kenyon, and
others. His comments have been cited by many authors, such as Menez (2018): “the
skull was discovered in the brecciated talus is therefore quite possible.” However,
he also added the he “not understand why Dr Busk should have considered that it
[the skull] was derived from the superficial part.” and “For the talus is, in fact,
exposed vertically throughout a very wide extent.”
While quoting
Duckworth precisely, some details were left out, which, add more context. I’ll
come back to that later.
Duckworth (1910),
also contemplated that the cave might be a possibility. Having assessed the
area of Forbes’ Quarry to be composed of “more solid rock”, (compared to the
brecciated talus), it would be “excluded at once, were it not that just at this
spot it contains a cave.” He explored the cave from 13 to 17 September, but
found it contained “nothing save the very earliest and seemingly marine
deposits covered with stalagmite.”
The answer to his
query as to why Busk had opined that the Gibraltar 1 skull came from the
“superficial” part of the talus had been staring him in the face as he crossed
the floor of the quarry: the lime kiln. Living in the Peak District, as I do, I
have often observed that those labourers of the 19th century, when
fetching limestone to ‘burn’ in their kilns, naturally start with the material
closest to the kiln. In the case of Forbes’ Quarry, the brecciated slope must
have, been seen, as an excellent source limestone for produced quicklime
(calcium oxide). Therefore, in the 50 years, between Busk and Falconer’s visit
to the quarry, the local lime burners had obviously worked the slope of Busk
and Falconer back into the vertical outcrop of Duckworth. Whatever the position
of the skull in the talus, the point was made moot as on Christmas Day 1910, an
absolutely massive, landslip almost filled the quarry entirely.
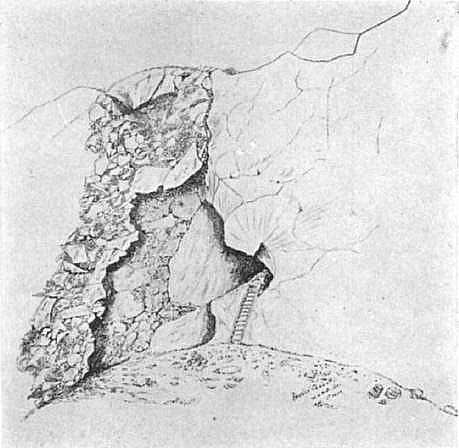
Duckworth’s (1911) sketch of the Forbes’ Quarry Cave. Duckwoth’s comments, verbatim, form the original paper: “The actual appearance of the surface exposed by the workings in this quarry can be described more clearly with the aid of the sketch (Plate XL, Fig. 1), to which reference will now be made. The face that has been worked must have had much the same character throughout and it is quite peculiar, for the quarry lies exactly at the zone of union of the solid rock, shown in Plate XL, Fig. 1, to the right, with an extraordinary mass of consolidated debris known as the "brecciated talus."
That the skull was
discovered in the brecciated talus is therefore quite possible, but I do not
understand why Dr. Busk should have considered that it was derived from the
superficial part. For the talus is, in fact, exposed vertically throughout a
very wide extent.” This drawing makes it clear that the brecciated talus, was
to the left of the cave, and that the removal of it, presumably for lime
burning, did indeed remove a great deal of it, with the workings moving right
to left. A modern photograph shows the scene today:

Menez (2018), photograph of the location of Forbes’ Quarry Cave. Remnants of brecciated talus can still be seen on the left of the pillbox and solid rock to right. This, seems to me, to point to an origin for Gibraltar 1, higher up the slope, in a cave which, has been eroded away. Original caption reads: “Figure 4. The location of the cave at Forbes’ Quarry in 2016 (author’s photograph). The cave is still accessible from within the Second World War pillbox.”
Breuil (1922) the
area three times over the next decade (1911, 1917 and 1919) and commented on
Forbes’ Quarry and examined “the foot of
the slopes formed by the rocky rubbish of Forbes Quarry.” He noted that the
breccia was no longer in situ “on account of a gigantic landslide”. He also noted
that the “old marine rock-shelter of Forbes Quarry [the cave] was of no
interest, and had never contained deposits other than marine gravels and a
layer of stalagmite and clay, with the bones of very small mammals.”
On his perambulations, around the area, Breuil did notice that an intact talus slope fronted Devil’s Tower Cave nearby. He was unable to excavate this himself, and recommended it to his student, Dorothy Garrod, as a likely place to seek further Neanderthal remains.
Now I must step back
many years to look at how archaeologists classified the Gibraltar 1 skull. The
first to give a detailed description was Broca (1868). He gives some
measurements and makes a few comparisons, which while not a formal diagnosis,
highlighted the differences between it and save but one other, fossil skulls
excavated up to 1868.
His description reads
thus:
“3° Another
prehistoric skull. The last two photographs sent to us by Mr. Busk represent
the face and profile of an extremely curious skull, which also comes from the
vicinity of Gibraltar, but which appears much older than the previous ones [from Genista cave]. The date of this
skull is also undetermined! It was not found in caves, but in the surrounding
soil. It was buried in a very compact, very adherent matrix, from which we
could only release with the greatest difficulty.
A large part of the
vault, between the bregma and the lambda, is absent. What remains of the vault
presents a slight degree of asymmetry which seems to be the consequence of a
posthumous deformation. I cannot give geological information on the ground from
which this skull was extracted. According, to Mr. Busk's communication to the
Norwich Congress, no characteristic fossils were found there; but everything
indicates, moreover, that this site is extremely old, and the skull itself
presents characteristics of inferiority, which fully confirm this view, and to
which Professor Huxley has drawn the attention of members of Congress.
The absence of part
of the vault and the fragments of gangue which still adhere to several points
do not allow the diameters to be rigorously measured in order to determine the
cephalic index, but it is evident that this skull is very dolichocephalic. It
is not very bulky; but its walls are very thick; around the perimeter, the
thickness of the parietals amounts to 9 and a half millimeters. The
superciliary arches form a considerable projection on the profile; the forehead
is small and very elusive.
The face is broad and
prognathous, the opening of the anterior nostrils is very wide, the eye sockets
are enormous and almost rounded in shape. Their width is 44 millimeters, their
height 39, their depth 51. The width of 44 millimeters is the largest I have
found so far on a human skull; it is exactly that of the orbits of the old man
of Les Eyzies, but in the latter case the enormous transverse development of
the orbital openings coincided with an excessive reduction of the vertical
diameter (27 millimeters), while on the Gibraltar skull the height of the orbit
is on the contrary exaggerated. As a result, despite the extreme width of the
orbit, the orbital index rises to 68.83, an enormous figure, nearly 4 percent
higher than the maximum that I have encountered so far in man. The interorbital
space is, moreover, very wide, 23 millimeters below and much larger above.
It follows that the transverse development of the upper part of the face is very great. On the front photograph, we do not see the temporal region, entirely masked by the outer edges of the eye sockets; the external orbital process forms a considerable protrusion, above which the forehead sharply narrows. This forehead is also extremely low, and it is so small in all its dimensions, especially when compared to the face, that it resembles that of apes. Professor Huxley has pointed out the simian shape of the dental arch, which tapers noticeably behind, like a horseshoe. The Company is already aware of the importance of this character, to which, for several years now, Mr. Alix has drawn its attention. Another simian character that Mr. Huxley insisted on is the absence of the canine fossa, which is replaced by a convex surface! Mr. Huxley has so far not seen this conformation on any other human skull, and I believe it can be said that it is not found on none of the skulls in our museum.”
The next scientist to
analyse the Gibraltar 1 skull was Sollas (1908). His work built on that by
Huxley (1863) and more especially Schwalbe (1890; 1901a; 1901b; 1904; 1906).
The extremely detailed work discusses the problems of measuring skulls
accurately including that of establishing a baseline from which to measure and
what equipment to use. Sollas measures, computes and compares many lengths,
radii and angles of curvature, with reference to a collection of Australian
skulls, Neanderthal 1, the Krapina remains, the skulls from Spy, more recent
humans and certain great apes. In short the work, running to 60 pages, with
copious illustrations and tables of data is a tour de force, the thoroughness
of which, many modern papers do not attain.
Sollas comments: “One
of the most important, as it is certainly one of the most striking, peculiarities
of the Neandertal calotte is the frontal torus, the confluent supra-orbital
tori; this is distinguished not merely by its magnitude, though this is
excessive, but still more, as Schwalbe points out, by the continuous and
uniform character which it maintains throughout its whole extent. In the skulls
of Australian natives two regions may usually, though not always, be
distinguished in each supra-orbital torus, a more or less shallow groove which
takes an oblique course, separating an
outer temporal from a
more median supra-ciliary region; in the Neandertal group this groove is
almost, if not entirely, effaced.” Sollas goes on to conclude: “Briefly
summarising our results, we may remark that the skull of the Neandertal race
possesses many features in common with certain flattened skulls which are met with
among tribes inhabiting the southern part of Australia ; it differs from them in
breadth, being markedly broader, in the characters of the glabellar region, and
in thickness. The face of the Neandertal race on the other hand is peculiar.
The large round, widely open orbits, the projecting broad nose, the retreating
cheek bones, the absence of any depression beneath the orbits, the long face,
and the low degree of prognathism distinguish it in the clearest manner from
the Australian. “
While Sollas (1908),
does not address the thorny issue of nomenclature, he has established two very
clear points:
1. Gibraltar 1.
Belongs to the ‘Neanderthal race’, since referred to Homo neanderthalensis.
2. More importantly, Sollas has established a system of measurement and comparison for human skulls and Neanderthals in particular.

Palate of Gibraltar 1 skull from Sollas (1908). Original caption reads: Fig. 24. – Outline of the palate of the Gibraltar skull, drawn with an orthopter. ( x 2/3.)
One last point regarding the actual circumstances of the discovery of Gibraltar 1 is given in Menez (2018): “It may be that an old lady who lived in the small fishing village of Catalan Bay, very near Forbes’ Quarry, was the last living person with first hand knowledge of the skull’s discovery. In his letter to Sir Arthur Keith of 28 April 1910, Samuel William Turner [Turner 1910], reported that: Today I drove round the Rock to Catalan Bay on the Eastern Side to interview an old woman of 84 who was said to know something about the finding of this skull, but I found the old lady almost in her dotage, and could not get reliable facts from her. The lady would have been 22 years old in 1848, and 38 during Busk and Falconer’s visit in 1864. Had she been known to them, we might have a much fuller history of the Gibraltar Skull.”
The information, or
rarther lack of it, that Turner gained from the old lady of Catalan Bay,
perhaps relates to the apocryphal story I presented at the beginning of this
piece? Whatever the true circumstances of the discovery of the Gibraltar 1
skull were, it is almost certain that we will never know.
References
Broca, P. (1869)
Remarques sur les ossements des cavernes de Gibraltar. In: Bulletins de la
Société d'anthropologie de Paris, II° Série. Tome 4. pp. 146-158;
Brown, A. (1867)
On the geology of Gibraltar, with especial reference to the recently explored
caves and bone breccia. Proceedings of the Royal Artillery Institution,
Woolwich 5: 295–304.
Busk, G. (1864a)
Pithecoid priscan Man from Gibraltar. The Reader (London) 4 (23 July): 109–110.
Busk, G. (1864b)
Ancient human cranium from Gibraltar. The Bath chronicle. Special daily edition
September 22 1864: p. 3.
Duckworth,
W. L. H. (1911). Cave exploration at Gibraltar in September, 1910. Journal of
the Royal Anthropological Institute of Great Britain and Ireland 41: 350–380.
Huxley T.
H. (1863). On Some Fossil Remains of Man, in
Evidence as to Man's place in nature. Williams & Norgate, London.
Murchison,
C (1868). Hugh Falconer, biographical sketch in Falconer Palaeontological Memoirs and Notes of the Late Hugh
Falconer, 1868.
London:
Spottiswoode and Co; 1868:xlix.
Mayer, F.
J. C. (1864a). Ueber die fossilen
Ueberreste eines menschlichen Schädels und Skeletes in einer Felsenhöhle des
Düssel- oder Neander-Thales. In: Archiv für Anatomie, Physiologie und
wissenschaftliche Medicin. (Müller's Archiv), Heft 1, 1864, S. 1–26.
Mayer, F.
J. C. (1864b). Zur Frage über das Alter und die Abstammung des
Menschengeschlechtes. In: Archiv für Anatomie, Physiologie und
wissenschaftliche Medicin. (Müller's Archiv), 1864, S. 696–728
Menez, A. (2018).
The Gibraltar Skull: early history, 1848–1868. Archives of natural history,
45(1), pp.92-110.
Pavid, K.
(2019). A new look at the Gibraltar Neanderthals. NHM at: https://www.nhm.ac.uk/discover/news/2019/july/a-new-look-at-the-gibraltar-neanderthals.html accessed 21/08/2021
Schrenk, F.
and Müller, S. (2010). Die Neandertaler C. H. Beck München.
Schwalbe, G. (1890). Studien iiber
Pithecanthropus erectus,” ‘Zeitschr. f. Morph, u. Anthropol.,’ v.1,
pp. 16-240.
Schwalbe,
G. (1901a). Der Neandertalschiidel,” ‘Bonner Jahrbiicher,’ part 106.
Schwalbe,
G. (1901b). Uber die specifischen Merkmale des Neandertalschadels,” ‘
Verhandlungen der anatom. Gesellsch.’ 15 Versammlung in Bonn, pp. 44-61.
Schwalbe,
G. (1904). Die Vorgesehichte des Menschen, Braunschweig.
Schwalbe,
G.A. (1906). Studien zur Vorgeschichte des Menschen (Vol. 1) p. 154 E. Nägele.
Sollas,
W.J., 1908. VII. On the cranial and facial characters of the neandertal race.
Philosophical Transactions of the Royal Society of London. Series B, Containing
Papers of a Biological Character, 199(251-261), pp.281-339.
Turner, S. W. (1910). Letters to Sir Arthur Keith, Archive of the Royal College of Surgeons of England, London: Folder KL II MS0018/1/16/1–19. Now held at the Natural History Museum

























{kind=link}